Issue:
110
Page: 44-49
Cannabis Taxonomy: The ‘Sativa’ Vs. ‘Indica’ Debate
by Robert C. Clarke, Mark D. Merlin, PhD
HerbalGram.
2016; American Botanical Council
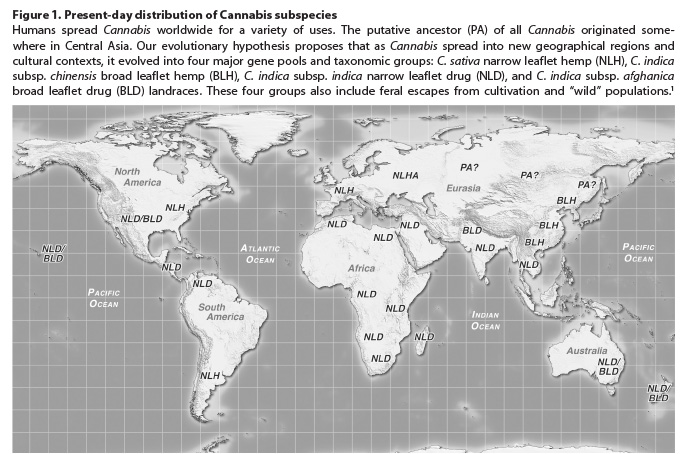
Marijuana and hemp (Cannabis) and the closely related
hop genus (Humulus) are the only widely known genera included in the
small, but economically valuable, Cannabaceae family. Swedish botanist Carl
Linnaeus, the “father of modern taxonomy,” first published the scientific name Cannabis
sativa in his seminal Species Plantarum of 1753. The Latin name
Cannabis derives from Greek (kannabis) and may have been originally
derived from Scythian. The term sativa simply means “cultivated” and
describes the common hemp plant that was widely grown across Europe in
Linnaeus’ time. We, the authors, consider C. sativa to be native to
western Eurasia and especially Europe, where, for millennia, the plant has been
grown for its strong fibers and nutritious seeds, and from where it was
introduced to the New World multiple times during early European colonization. Cannabis
sativa plants also produce very small amounts of the compound
delta-9-tetrahydrocannabinol (THC), the medically valuable and primary
psychoactive cannabinoid found only in Cannabis. Since C. sativa
evolved within the geographical limits of western Eurasia, it represents only a
small portion of the genetic diversity seen in the genus Cannabis
worldwide.1
In 1785,
European naturalist Jean-Baptiste Lamarck described and named a second species,
Cannabis indica, meaning “Cannabis from India,” after the origin of
the first samples of this highly psychoactive plant that reached Europe. Cannabis
indica has the genetic potential to produce relatively large amounts of
THC. The species is used for marijuana and hashish production, but in many
regions of eastern Asia it also has a long history of cultivation for fiber and
seed. Humans make cloth out of C. indica fibers and eat the seeds, but
this native eastern Eurasian species is more commonly used today as a drug
plant with widespread social and medicinal importance reaching well beyond its
original geographical range.1
While Karl
Hillig, PhD, was a doctoral student at Indiana University, he used
morphological and chemical characteristics to investigate the diversity of the Cannabis
genus and proposed taxonomic groupings (subspecies) that support the
original two-species concept.2-5 Hillig recognized European
cultivated Cannabis as a separate species (C. sativa). Because
this species typically has narrow leaflets and is primarily used for hemp fiber
and seed production, we refer to it as narrow leaflet hemp (NLH). European C.
sativa NLH populations are much less genetically diverse than those found
in many other regions.
Hillig
assigned the remainder of the world’s cultivated varieties to C. indica
and divided them into three subspecies. One of these subspecies, C. indica
subsp. indica, includes varieties that span the Indian subcontinent from
Southeast Asia to western India and into Africa. These traditional drug
varieties produce abundant amounts of THC with little if any cannabidiol (CBD).
CBD is the second most common cannabinoid, and is non-psychoactive, but it has
been shown to be medicinally effective for a variety of indications. By the
19th century, high-THC C. indica subsp. indica reached the
Caribbean region and steadily spread throughout Central and South America. Since
the 1960s, most of the drug Cannabis that reached North America and
Europe was C. indica subsp. indica. Marijuana users commonly call
domestically grown plants of these varieties “sativas” because their leaflets
are relatively narrow, and therefore exhibit a superficial resemblance to those
of European NLH plants. However, in our construct, this is a misnomer as C.
sativa plants produce little if any THC. Based on Hillig’s research, we now
refer to members of C. indica subsp. indica as narrow leaflet drug
(NLD) varieties. Although they have relatively narrow leaflets like NLH (C.
sativa) plants, the NLD plants can produce an abundance of THC and are most
commonly used for their psychoactive effects. Based on taxonomic tradition,
these plants are properly called “indicas” rather than “sativas.”
A second C.
indica subspecies originated in Afghanistan where crops were traditionally
grown to manufacture sieved hashish, a mechanically concentrated Cannabis
drug. From 1974, when descriptions and photos of Afghan Cannabis
were published by Harvard professor Richard E. Schultes, PhD, it became readily
apparent that it represented a type of drug Cannabis previously unknown
outside of Eurasia, belonging neither to Linnaeus’ C. sativa nor
Lamarck’s C. indica.6 Its shorter, more robust stature, and
broad, dark green leaves easily distinguish it from the taller, lighter green,
and more laxly branched NLD varieties. Because of its limited geographic range
and restricted usage, the Afghan genome is less diverse than the NLD genome. By
the late 1970s, seeds of Afghan hashish varieties reached Europe and North
America and were rapidly disseminated among marijuana growers. At this time,
all Cannabis varieties were commonly considered to be members of a
single species, C. sativa, and the familiar NLD marijuana varieties were
called “sativas” to differentiate them from the newly introduced and quite
different looking Afghan varieties commonly called “indicas.” Hillig named them
C. indica subsp. afghanica, which we now refer to as broad
leaflet drug (BLD) varieties to differentiate them from NLD varieties. On
average, populations of BLD plants contain approximately equal amounts of THC
and CBD. Although BLD varieties are also considered by us to be members of C.
indica, it is more correct to distinguish them from subspecies indica
from India by calling them subspecies afghanica, or simply “Afghans.”
Hillig’s third
grouping within C. indica is subspecies chinensis, which
comprises the traditional East Asian fiber and seed varieties and associated
feral populations. We refer to this group as broad leaflet hemp (BLH). Like
other subspecies of C. indica, varieties of C. indica subsp.
chinensis possess the genetic potential to produce psychoactive THC, but
East Asian cultural traditions, such as Confucianism, have long encouraged the
selection of these varieties for their economically valuable fiber and seed,
rather than their psychoactive potential. As a result, total cannabinoid
production is lower than in subspecies indica and afghanica.

Evolutionary
theory predicts that, at some point in time, there must have been a putative
ancestor of the two modern species, C. sativa and C. indica. This
ancient ancestor is often referred to as C. ruderalis, which may have
originated somewhere in Central Asia. However, by now it is probably extinct,
and seemingly ancestral populations are more likely descendants of feral plants
that escaped from cultivation long ago. Evolutionary hypotheses based on plant
distribution studies, paleoclimate modeling, archaeological evidence, and the
historical record propose that C. sativa NLH most likely originated in a
temperate region of western Eurasia, possibly in the foothills of the Caucasus
Mountains, from a putative hemp ancestor with diminished biosynthetic potential
to produce THC. Cannabis indica likely originated in the Hengduan
Mountains, in present-day southwestern China, from a putative drug ancestor
that had evolved an enhanced ability to produce THC. Early C. indica
populations diversified as they were introduced by humans to different
geographical regions where they may have further evolved into the three
subspecies, all of which produce THC.1
Cultivated
plant varieties are called cultivars, and when cultivars are grown and
maintained by local farmers over generations, we refer to them as landrace
cultivars, or landraces. Landraces evolve in a balance between natural
selective pressures exerted by the local environment favoring survival, and
human selections favoring a cultivar’s ability to both thrive under cultivation
and produce particular culturally preferred products. Early humans spread Cannabis
into many new regions as they moved, and at each new camp or settlement they
selected seed from superior plants that were suited to their own uses and
processing methods. By sowing seeds from the most favorable individuals,
traditional farmers developed and maintained the landraces upon which
present-day hybrid hemp and drug cultivars were founded.
Hemp cultivars
were derived from crosses between different European NLH landraces and East
Asian BLH landraces. Traditional Asian, African, and New World drug
landraces were, until relatively recently, all pure NLD types. Before the
introduction of BLD landraces from Afghanistan in the late 1970s, hybrids
between imported NLD landraces formed the core genome of domestically produced
drug Cannabis in both North America and Europe. It is through crossing
NLD and BLD landraces from such geographically isolated populations that modern
hybrid sinsemilla (Spanish for “seedless”) cultivars were created.1
Unfortunately,
we cannot return today to a region previously known for its fine Cannabis
and expect to find the same landraces that were growing there decades before. Cannabis
is open-pollinated, with male and female flowers borne on separate plants, and,
therefore, two plants are usually required to produce a seed. Random
combinations of alleles (forms of a gene) and accompanying variation are to be
expected. Cannabis landrace varieties are best maintained by repeated
natural and human selection in situ — nature selecting for survival and
humans selecting for beneficial traits. Without persistent human selection and
maintenance, these landrace varieties will tend to drift back to their
atavistic, naturally selected survival mode.
The Western
world began using imported marijuana and hashish in the 1960s, and all of the
remarkable imported varieties available then were traditionally maintained
landraces. Within a decade, the demand for quality drug Cannabis
exceeded traditional supplies, and mass production in the absence of selection
became the rule. Rather than planting only select seeds, farmers began to sow
all their seeds in an effort to supply market demand, and the quality of
commercially available drug Cannabis began to decline. In addition,
travelers returned to the supplying nations and introduced seeds of “improved”
Western sinsemilla varieties that interbred with the local landraces and
thus contaminated the local genomes. Landraces can no longer be replaced; they
can only be preserved. The few remaining pure landrace varieties in existence
now, some kept alive for decades as seeds and cuttings, are the keys to future
developments in drug Cannabis breeding and evolution. It will be a
continuing shame to lose the best results of hundreds of years of selection by
local farmers. After all, our role should be as caretakers preserving the
legacy of traditional farmers for the future benefit of all.
Cannabis research is a work in progress, and
not all researchers agree on a single taxonomy.7 DNA sequencing is
currently being used to characterize the diversity of many plant and animal
groups, including Cannabis. While our knowledge grows and the
evolutionary history of Cannabis is revealed, changes in taxonomic
nomenclature will continue to reflect our deepening understanding of this
medically valuable, yet controversial, plant. More broadly, whether we discover
that Cannabis plants belong to one or more species, we can be sure that
humans have long known, used, dispersed, cultivated, and artificially selected
these plants to perpetuate a truly wide range of diversity.
Robert C.
Clarke is the
author of several Cannabis science books and has traveled extensively
throughout Eurasia documenting traditional Cannabis production and use.
His breeding interests include selection and preservation of landrace
varieties, and developing narrow leaflet drug varieties and hashish cultivars.
Clarke is the co-founder and director of BioAgronomics Group, an international
cannabis industry consultancy, serves as projects manager for the International
Hemp Association, and holds a seat on the Phylos Bioscience Cannabis
Evolution Project scientific advisory board. He may be contacted at rob@bioagronomics.com.
Mark D.
Merlin, PhD, is a
professor in the botany department of the University of Hawai`i at Mānoa and an
elected member of the Linnean Society of London. A large part of his long-term
biological and historical research has involved drug plants and their past and
present uses by people. He has authored or co-authored a number of books on
this general subject, including Man and Marijuana: Some Aspects of their
Ancient Relationships (Fairleigh Dickinson University Press, 1972), On
the Trail of the Ancient Opium Poppy (Associated University Presses, 1984),
Kava: The Pacific Drug (co-authored with Vincent Lebot and Lamont
Lindstrom; Yale University Press, 1992), and, most recently, Cannabis:
Evolution and Ethnobotany (co-authored with Robert C. Clarke; University of
California Press, 2013). He may be contacted at merlin@hawaii.edu.
References
- Clarke RC, Merlin MD. Cannabis: Evolution and Ethnobotany. Berkeley, CA: University of California Press; 2013.
- Hillig KW. A chemotaxonomic analysis of terpenoid variation in Cannabis. Biochem Syst Ecol. 2004;32(10):875-891.
- Hillig KW. A multivariate analysis of allozyme variation in 93 Cannabis accessions from the VIR Germplasm Collection. J Indust Hemp. 2004;9(2):5-22.
- Hillig KW. Genetic evidence for speciation in Cannabis (Cannabaceae). Genet Resour Crop Ev. 2005;52(2):161-180.
- Hillig KW, Mahlberg PG. A chemotaxonomic analysis of cannabinoid variation in Cannabis (Cannabaceae). Am J Bot. 2004;91(6):966-975.
- Schultes RE, Klein WM, Plowman T, Lockwood TE. Cannabis: An example of taxonomic neglect. Bot Mus Leafl Harv Univ. 1974;23(9):337-364.
- Small E. Evolution and classification of Cannabis sativa (marijuana, hemp) in relation to human utilization. Bot Rev. 2015;81(3):189-294.
|